Circuits de neurones impliqués dans la vision fovéale
Contexte
Si, chez les oiseaux, la perception d’objets en mouvement – vitesse, trajectoire et direction – dans un environnement complexe garantit leur survie, on ignore presque tout des circuits neuronaux impliqués dans la vision à haute fréquence et à haute résolution.
Dans la fovéa humaine et, plus généralement dans celle des primates, la voie dite midget, constituée d’un cône connecté à une cellule bipolaire (CB), qui est elle-même connectée à une seule cellule ganglionnaire (CG) dite naine, est le circuit neuronal dominant. Ce circuit est idéal pour une acuité spatiale élevée, mais demeure peu adapté à la perception du mouvement. La fovéa des primates est d’ailleurs peu performante dans le traitement de signaux visuels de fréquence élevée. Cette faible acuité spatio-temporelle au niveau de la fovéa est la principale raison pour laquelle nous percevons des images en mouvement plutôt qu’une séquence d’images séparées lorsque l’on regarde un film au cinéma ou sur notre smartphone.


La perception spatio-temporelle élevée des oiseaux modernes (Neoaves) suggère que leur rétine et les régions de leur cerveau impliquées dans le traitement des signaux visuels sont dotés de propriétés particulières.
Des expériences comportementales visant à estimer l’acuité visuelle temporelle ont montré que certains oiseaux peuvent distinguer des cycles alternés lumière-obscurité jusqu’à 145 Hz, soit > 50 Hz au-dessus de la fréquence détectée chez l’être humain (Bostrom et al., 2016 ; Potier et al., 2020)
Une question qui se pose est de déterminer si les oiseaux modernes déploient dans leur fovéa des circuits de neurones qui leur permettent de détecter avec netteté des objets (ex: insectes) en déplacement rapide.
Approche
Deux voies parallèles de transmission des signaux lumineux pourraient co-exister dans la fovéa des oiseaux :
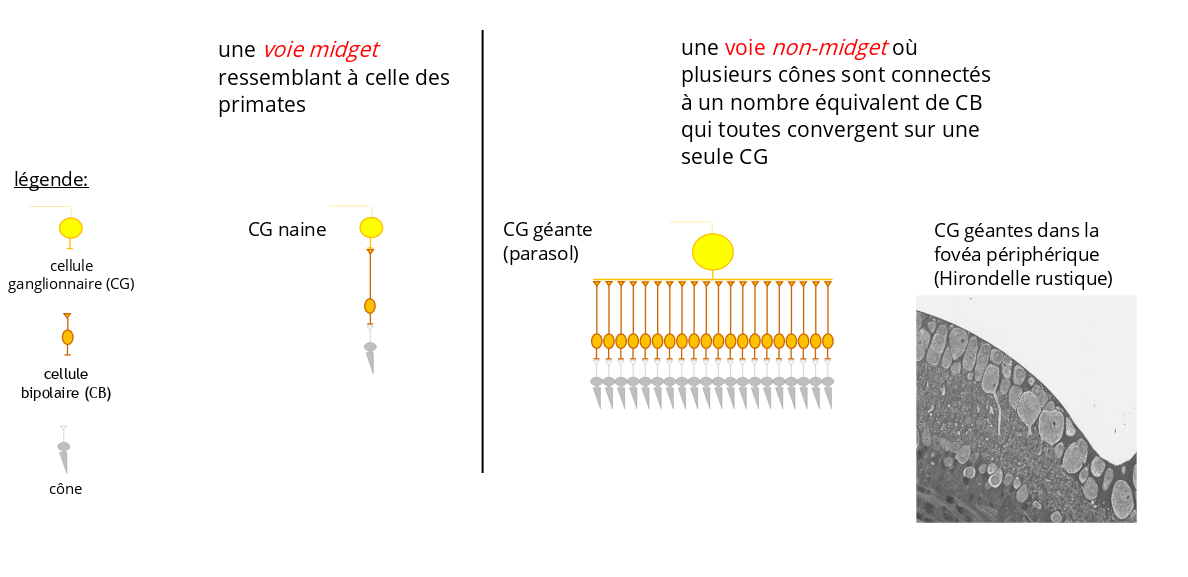
Voies midget et non-midget dans les régions de hautes acuités de la rétine
Les indices suggérant que les voies midget et non-midget co-existent dans la fovéa des oiseaux sont présentés dans l’article de Rodrigues et al. (2022) : https://www.biorxiv.org/content/10.1101/2022.10.29.514341v1.
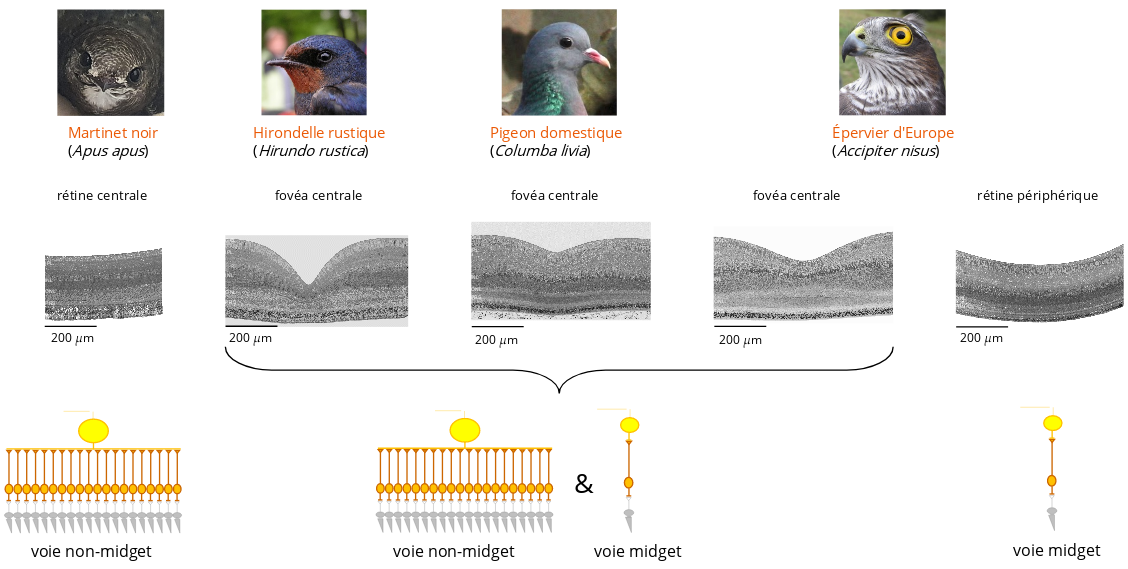
La présence de voies non-midget aurait l’avantage de fournir une large gamme de sensibilités aux variations rapides de stimulation lumineuse et offrirait ainsi à la fovéa des conditions nécessaires pour une perception à haute résolution des mouvements rapides.
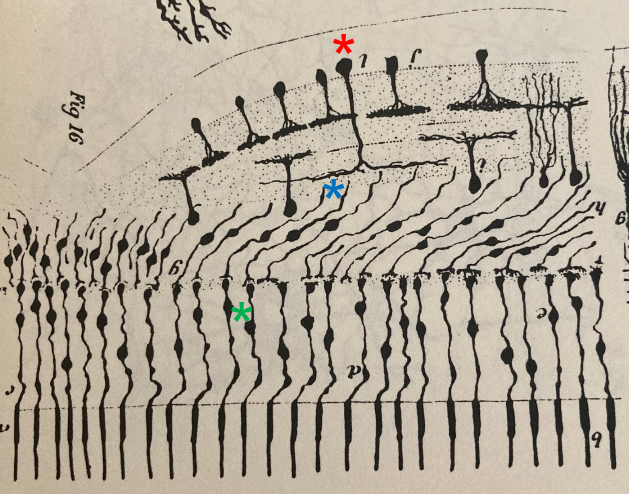
Santiago Ramon y Cajal (1852-1934) – le père fondateur des Neurosciences – avait déjà anticipé la présence d’une voie non-midget dans la fovéa des oiseaux. Un dessin de sa main illustre comment l’arbre dendritique d’une CG parasol (*) forme des connections avec plusieurs CB (*), chacune d’elle étant connectée à un seul cône (*) :
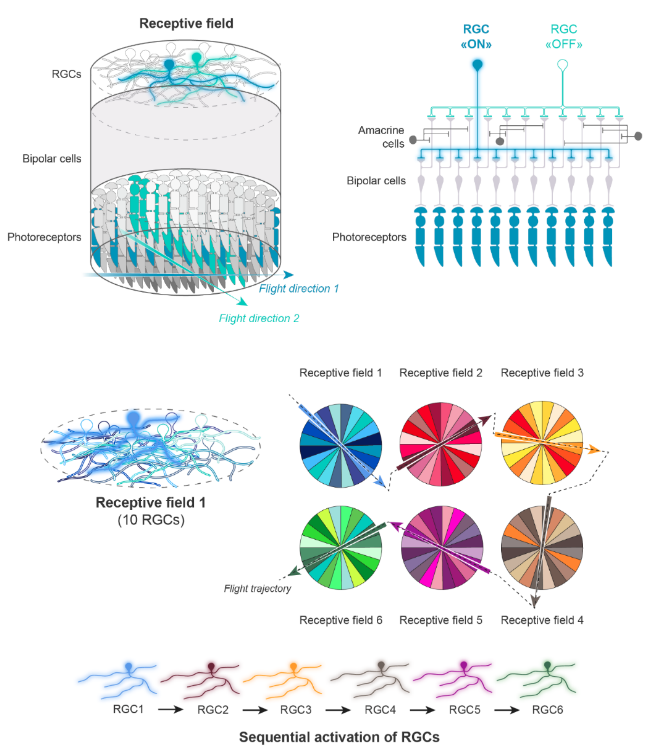
Modèle conceptuel d’organisation de la voie non-midget pour détecter la direction d’un objet en mouvement rapide
Dans ce modèle, un grand nombre de cônes appartenant à un même champ réceptif sont connectés, via des CB, à un petit nombre (dix dans notre exemple) de CG (dénotées RGC sur la figure). Ces CG ont comme caractéristique commune d’avoir des arbres dendritiques déployés dans la fovéa.
Les dendrites de ces dix CG sont entremêlées et définissent un seul champ réceptif de forme circulaire. Dans notre exemple, une CG parmi les dix (celle de couleur bleu clair dans les deux panneaux du haut et dans le champ réceptif 1) est activée (« ON ») par un objet en mouvement, comme un insecte volant. Elle l’est car les photorécepteurs qui activent cette CG définissent un domaine excitateur de forme allongée, qui a ici la même orientation que le signal lumineux fugace généré par l’insecte traversant le champ réceptif 1. Les neuf autres CG comme, par exemple, celle de couleur turquoise représentée dans les deux panneaux supérieurs et dans le champ réceptif 1, restent « OFF » car elles sont connectées, via des CB, à des photorécepteurs qui définissent des domaines excitateurs orientés selon des axes différents de celui qui correspond à la trajectoire de l’insecte. L’effet inhibiteur des cellules amacrines (CA) sur les CB (représentées dans le panneau supérieur droit) permettrait d’empêcher que la CG « turquoise» soit activée dans ces conditions. En se déplaçant à travers de nombreux champs réceptifs (six sont présentés ici), l’objet volant activerait séquentiellement les CG dont les axes d’orientation correspondent à sa trajectoire. Ce serait ainsi que la trajectoire de l’insecte serait « capturée » par la rétine.
Identifier et visualiser les neurones de la fovéa
Expériences réalisées en collaboration avec le laboratoire du Dr. Jean Livet :
https://www.institut-vision.org/fr/neurogenese-et-developpement-des-circuits-neuronaux.html
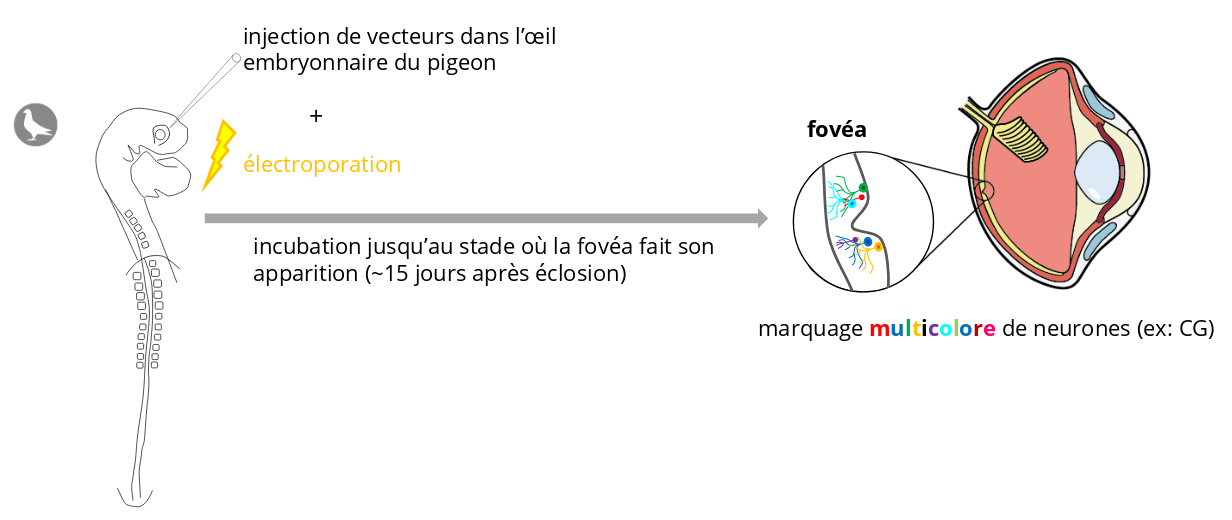
Cette expérience devrait nous apporter des éléments de réponse aux questions suivantes :
- Dans quelle proportion les CG de types parasol et naine sont-elles présentes dans la fovéa ?
- La visualisation dans un espace 3D des neurones fluorescents permet-elle de définir la place qu’occupent les arbres dendritiques des CG dans la fovéa ?
- Les arbres dendritiques des CG occupent-ils des espaces 3D qui se chevauchent ?
Remerciements
Ce projet est rendu possible grâce à la collaboration de François Jardin, colombophile et éleveur :
